Cell News 3/2013
19
RESEARCH NEWS
considered an emergent behaviour
7
(Figure 1d). In physics emer-
gent behaviours are commonly studied to describe complex
systems. Physics thus provides a framework for relating the mi-
croscopic properties of individual molecules to the macroscopic
properties of materials. This exactly is the key challenge in mo-
dern cell biology, bridging the gap between individual molecules
and their collective behaviour (Figure 1e).
A Multi-Scale Model for
Xenopus
Spindles Length.
During early
Xenopus
development cell size decreases dramati-
cally from a 1200 µm diameter fertilized egg to approximate-
ly 12 µm diameter blastomeres
8
. Consequently, the metaphase
spindle needs to function in cell volumes that vary by several
orders of magnitude. Because defects in spindle length result in
erroneous cell division
9
, robust mechanisms to set the length of
a spindle and scale it according to cell type must exist. Deca-
des of work have shown that microtubule dynamics, nucleation,
and transport are critical for spindle assembly
10
. However, the
combined activities of these processes, as well as their complex
interplay, make it difficult to determine how each individual pro-
cess contributes to the overall organization of the spindle.
We have recently shown that mass balance together with a “li-
quid crystal” analogy is a useful working hypothesis to account
for the dynamic properties of
Xenopus
spindles and link these to
its overall geometry (Figure 2a). Mass balance, which is based
on the principle of mass conservation, is often used in physics
and engineering to infer properties of systems that are difficult
to measure directly. In the 1940s, Irvin Isenberg suggested that
the spindle might be a liquid crystal
11
. Indeed, more recent ob-
servations in
Xenopus
egg extracts imply that spindles can dis-
play liquid-like properties. For example, two meiotic spindles
fuse when brought close together
12
. Furthermore, micromanipu-
lation studies on
Xenopus
spindles show that spindles recover
their original shape after weak compressions and that they have
anisotropic material properties
13-16
. This coarse-grained “liquid
crystal” analogy has important implications, because spindle
length follows from a simple book-keeping exercise based on
mass balance: The length of the spindle will simply depend on
the balance of material that is created (nucleation and assembly)
and lost (disassembly), while the density of microtubules remains
roughly constant. At steady-state, the rate of volume increase
due to incorporation of tubulin into microtubules equals the rate
of volume loss due to microtubule disassembly. Consequently,
spindle length is ultimately related to the kinetic parameters of
microtubule nucleation, assembly and disassembly
17
(Figure 2b).
A key prediction of our work is that spindle length scales linear-
ly with microtubule growth velocity when other parameters of
spindle organization remain constant
17
. This prediction can be
tested in quantitative experiments that allow the modulation of
microtubule dynamics in
Xenopus
egg extract spindles. But how
can we specifically modulate microtubule growth rates? Work
over the last decade established a major role for XMAP215 in
microtubule growth promotion. XMAP215 was shown to act as a
processive microtubule polymerase that promotes incorporation
of tubulin into the growing plus end (Figure 3a). XMAP215 binds
the tubulin heterodimer in a 1:1 stoichiometry, ‘surfs’ the gro-
wing microtubule plus end, and stays there for multiple rounds
of tubulin incorporation
18
. XMAP215 was first isolated from
Xe-
nopus
eggs
19
and subsequently found in all major kingdoms of
eukaryotes, including fungi (Stu2
20
in
S. cerevisiae
, Dis1
21
and
Alp14
22
in
S. pombe
), plants (Mor1
23
in
A. thaliana
) and animals
(Zyg9
24
in
C. elegans
, mini spindles
25
in
D. melanogaster
, and
ch-TOG
26
in humans). All members of the XMAP215 family are
characterized by a varying number of TOG domains at their N-
termini (Figure 3b). However, how these various domains contri-
bute to XMAP215 activity was until recently not known. Based
on mutants in TOG domains that interfere with tubulin binding
27
,
it has been proposed that TOG binding to tubulin is required for
its catalytic activity
28
; however, there was no proof for this idea.
Recently, we have shown that the polymerase activity of
XMAP215 depends on tubulin binding to multiple TOG domains.
Mutation of conserved residues in different TOG domains redu-
ces tubulin binding while concomitantly reducing the maximal
growth promoting activity of the polymerase (Figure 3c). Com-
bining mutations in different TOG domains allowed us to mo-
dulate the enzymatic activity of XMAP215 and thereby control
microtubule growth velocity
in vitro
29
. This set of well-charac-
terized polymerase mutants served as an experimental tool kit
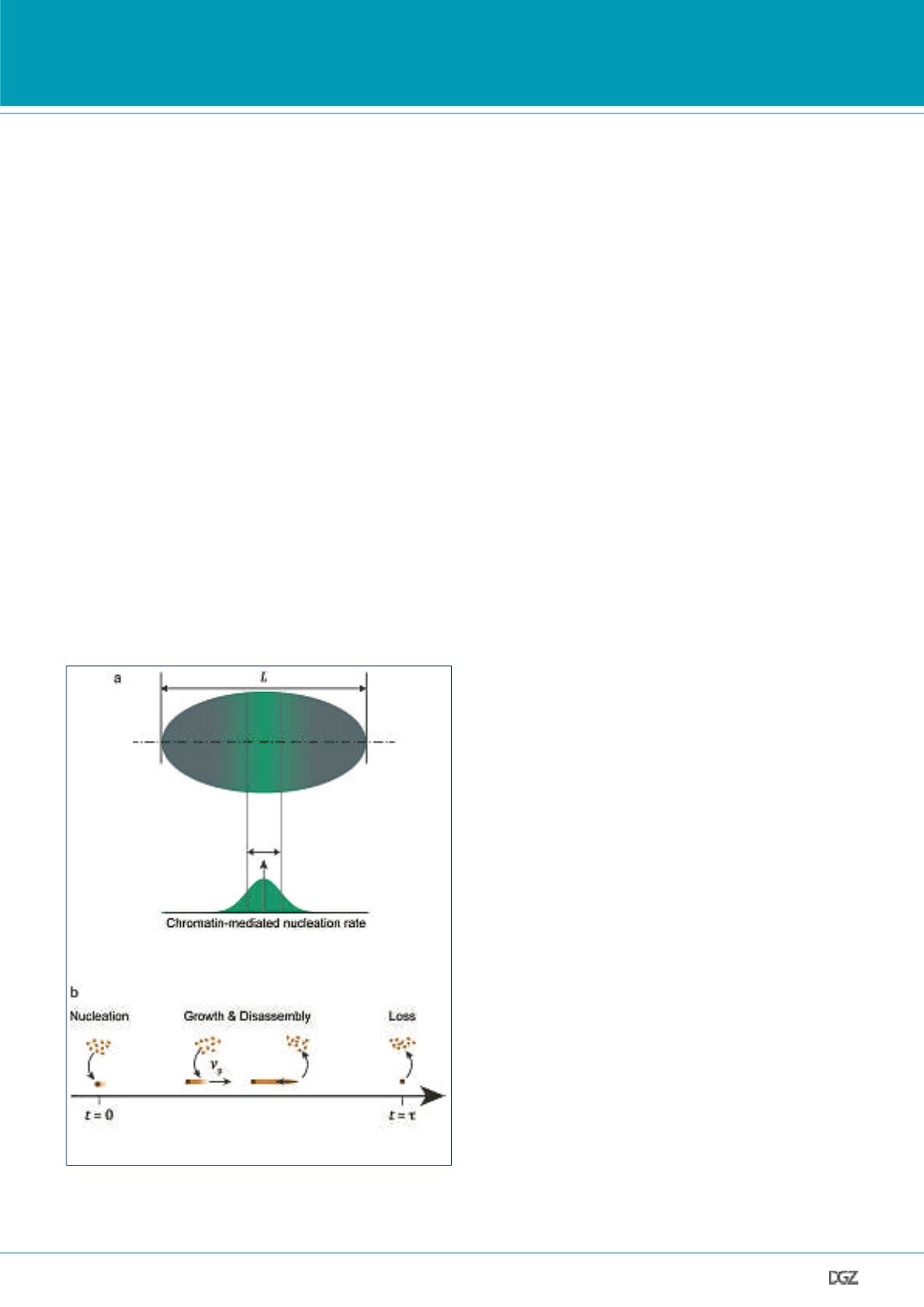
Figure 2. Xenopus Spindle Length:
(a) Mass balance together with a “liquid crystal” analogy allows us to rela-
te kinetic parameters of microtubule nucleation, assembly and disassembly
(b) and the overall geometry of Xenopus spindles.


