Cell News 4/2014
21
Research news
from a different cellular origin than papillomas (Brown et al.
1998; Perez-Losada and Balmain 2003) and are frequently ob-
served in humans (Burnworth et al. 2006). KAs may either arise
sporadically or as adverse effect e.g. upon BRaf inhibitor therapy
in melanoma patients probably due to paradoxical ERK activa-
tion downstream of endogenous CRaf (Oberholzer et al. 2012).
Interestingly, such CRaf activation also occurs in Par3-deficient
KAs (Iden et al. 2012b), suggesting that Par3 usually serves to
restrict CRaf function. Whereas the formation of papillomas
correlates with robust Par3 expression and localization to cell-
cell contacts, we found Par3 strongly reduced in murine and
human KAs and almost exclusively localized in the cytoplasm, in
line with tumor-suppressive properties of Par3 in KA.
Our results thus identified a dual function of Par3 in skin can-
cers of clinical relevance, with both (pro)oncogenic and tumor-
suppressive activity depending on the tumor type (Iden et al.,
2012b), and indicate that differential subcellular localization of
Par3 in distinct epidermal cell populations is an important para-
meter for tumor outcome.
Par3 acts as invasion suppressor in epidermal cancers
Disturbed polarity may also affect subsequent events of tumor
cell invasion and metastasis. Overexpression and cytoplasmic
accumulation of aPKC
λ/ι
is frequently associated with progres-
sion of different human cancers such as pancreatic, cervical,
breast cancer and hepatocellular carcinoma and poor survival
(Scotti et al. 2010; Huang and Muthuswamy 2010; McCaffrey
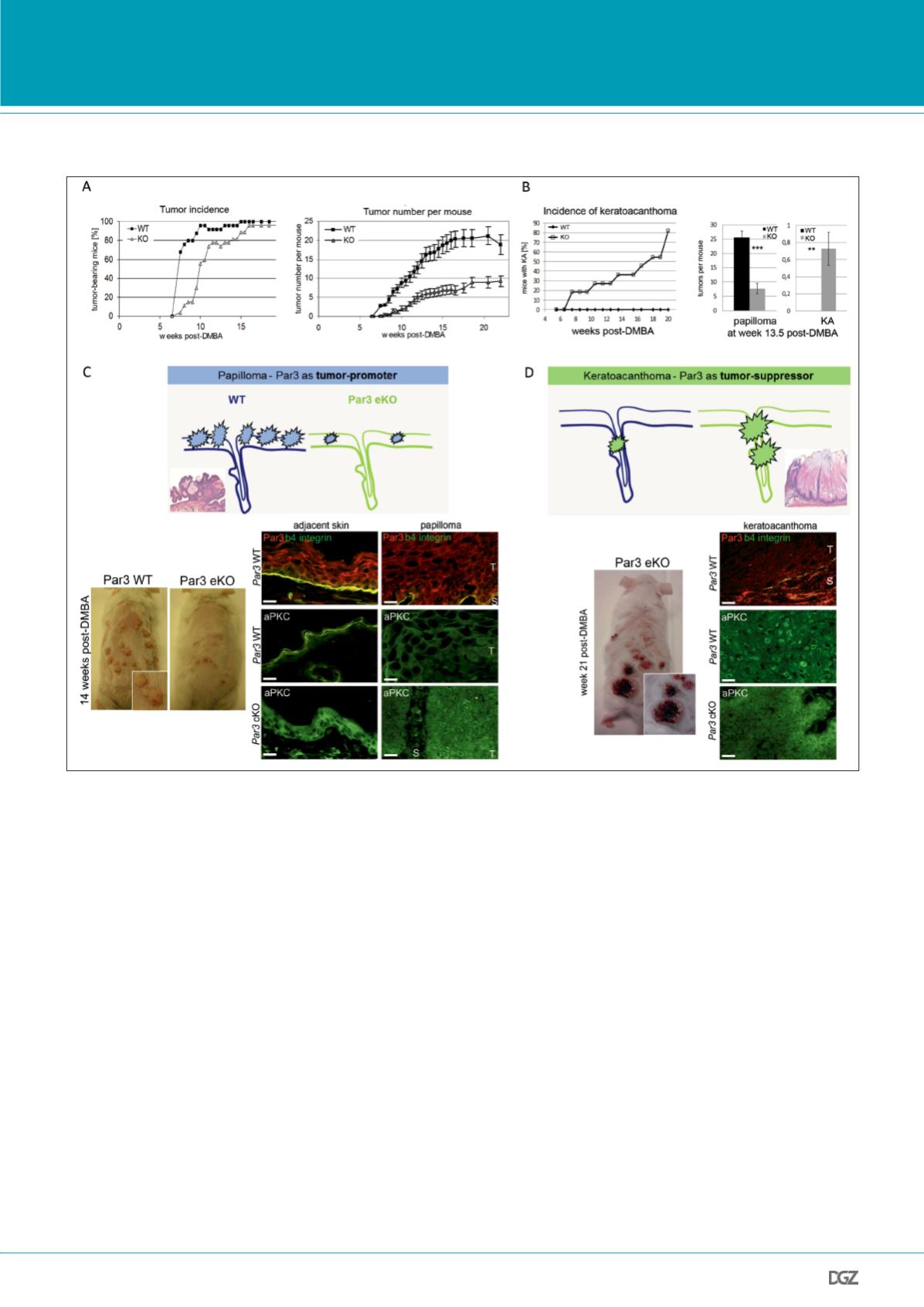
Figure 3: Dual role of the polarity protein Par3 in skin tumorigenesis.
A, left graph: Tumor incidence, indicated by percentage of tumor-bearing mice, is reduced in epidermal Par3 KO mice. n(WT) = 25, n(Par3 eKO) = 27. Right
graph: Average number of tumors per mouse is strongly reduced upon epidermal Par3 deletion (mean ± SEM). B, left graph: KA incidence based on macro-
scopic analysis of control and Par3 eKO mice demonstrated predisposition of Par3 eKO mice to this tumor type. Right panel: Number of papillomas versus
KAs per mouse based on macroscopic analysis at week 13.5 post-DMBA. C, top panel: model of tumor-promoting function of Par3 in papillomagenesis,
bottom panel: representative mice during DMBA/TPA skin carcinogenesis (14 weeks post-DMBA) illustrating reduced tumor number and size upon loss of
epidermal Par3, and Par3 and aPKC immunostaining in cryosections of mouse epidermis and papilloma. b4 integrin signal reflects the epidermal-dermal
border. D, top panel: model of tumor-suppressive function of Par3 in KA formation, bottom panel: representative Par3 eKO mouse during DMBA/TPA skin
carcinogenesis (21 weeks post-DMBA) illustrating typical KA, and loss of Par3 immunostaining as well as mislocalization of aPKC in cryosections of mouse
keratoacanthoma. Scale bars, 20 um. T, tumor; S, stroma. (modified from Iden et al., 2012b).


