Cell News 04/2019
11
PRIZE WINNERS 2019
thereby destabilizing junctional cadherins (i.e. AJs) and enabling
cells to delaminate during EMT (summarized in Figure 4).
Enhanced centrosomal MT dynamics control the
time of BP residence in the SVZ
We aimed to understand the function of Akna and MT dynam-
ics in SVZ-cells (where it is highest expressed) and the reason
for why it is downregulated in neurons. We observed that, in
contrast to neural progenitors (NSCs and BPs), neurons have a
greater degree of non-centrosomal MT growth. This means that
young neurons re-organize their MT cytoskeleton when they
mature and leave the SVZ towards the CP. We, therefore, chal-
lenged this process by counteracting the endogenous downregu-
lation of Akna through overexpression in neurons. While control
cells reached the CP, Akna overexpression inhibited migration,
with cells accumulating and differentiating at the SVZ and IZ
(Figure 5b). Importantly, this was not observed upon expression
of the non-centrosomal construct, again indicating that the cen-
trosomal localization is important. Hence, Akna maintains cells
within the SVZ and endogenous downregulation is a necessary
step to leave it. It is known that neurons inactivate centrosomes
as the main MT organizing center (MTOC) as shown by others
(Stiess et al., 2010) and here by us, and that neuronal MTs are
less dynamic and more stable (more detyrosination and acetyl-
ation) (Figure 5b). Our results indicate that Akna counteracts
these neuronal hallmarks by promoting centrosomal microtubule
nucleation, faster microtubule growth, and more dynamic and
less detyrosinated microtubules.
At which step are Akna levels critical for the transition of mul-
tipolar SZV cells to bipolar neurons migrating into the CP? Life
imaging of cortical slices showed that cells in which Akna was
prematurely downregulated transitioned faster to the bipolar
morphology, while keeping Akna protein levels high made them
retain a multipolar morphology and migrate less frequently. So,
the switch to bipolar morphology and radial neuronal migration
requires a switch from a centrosomal to more non-centrosomal
MTOC, and a reduction of MT dynamics, via endogenous down-
regulation of Akna (summarized in Figure 4)
Far-reaching implications
Our discoveries raise important questions in cell, developmen-
tal and cancer biology. Do MT dynamics have a conserved role
in phylogeny of the cerebral cortex? We analyzed the levels of
Akna (RNA and protein) in cells of the developing cerebral cortex
of ferrets and macaques, and in cerebral organoids derived from
human induced pluripotent stem cells, all of which have a folded
brain. Interestingly, we observed that Akna is highly expressed in
the expanded SVZ at the peaks of neurogenesis in these species,
and functional analyses in cerebral organoids offered concrete
evidence that the role of Akna in delamination of NSCs is con-
served during phylogeny.
Is Akna relevant for cell migration and polarization in other
tissues, and is this encoded in a genetic program? The transcrip-
tion factors Sox4 and Tcf12 are positive upstream regulators of
Akna gene expression (see summary in Figure 4), and both are
essential for delamination of NSCs and BP generation (Chen
et al., 2015; Mesman et al., 2017; Uittenbogaard et al., 2002).
Interestingly, both TFs are also required during delamination of
epithelial cells during EMT (Lee et al., 2012; Tiwari et al., 2013),
hence indicating that the cell’s ability to adjust the behavior
of its MTs is genetically encoded and can be regulated by a
network of TFs, which is indeed a logic way to link and tune the
architecture of the cytoskeleton with changes in cell identity.
Of notice, Akna levels are high in tumor tissue compared to
healthy specimens. This is particularly relevant in the context of
anti-cancer treatment development because it would suggest
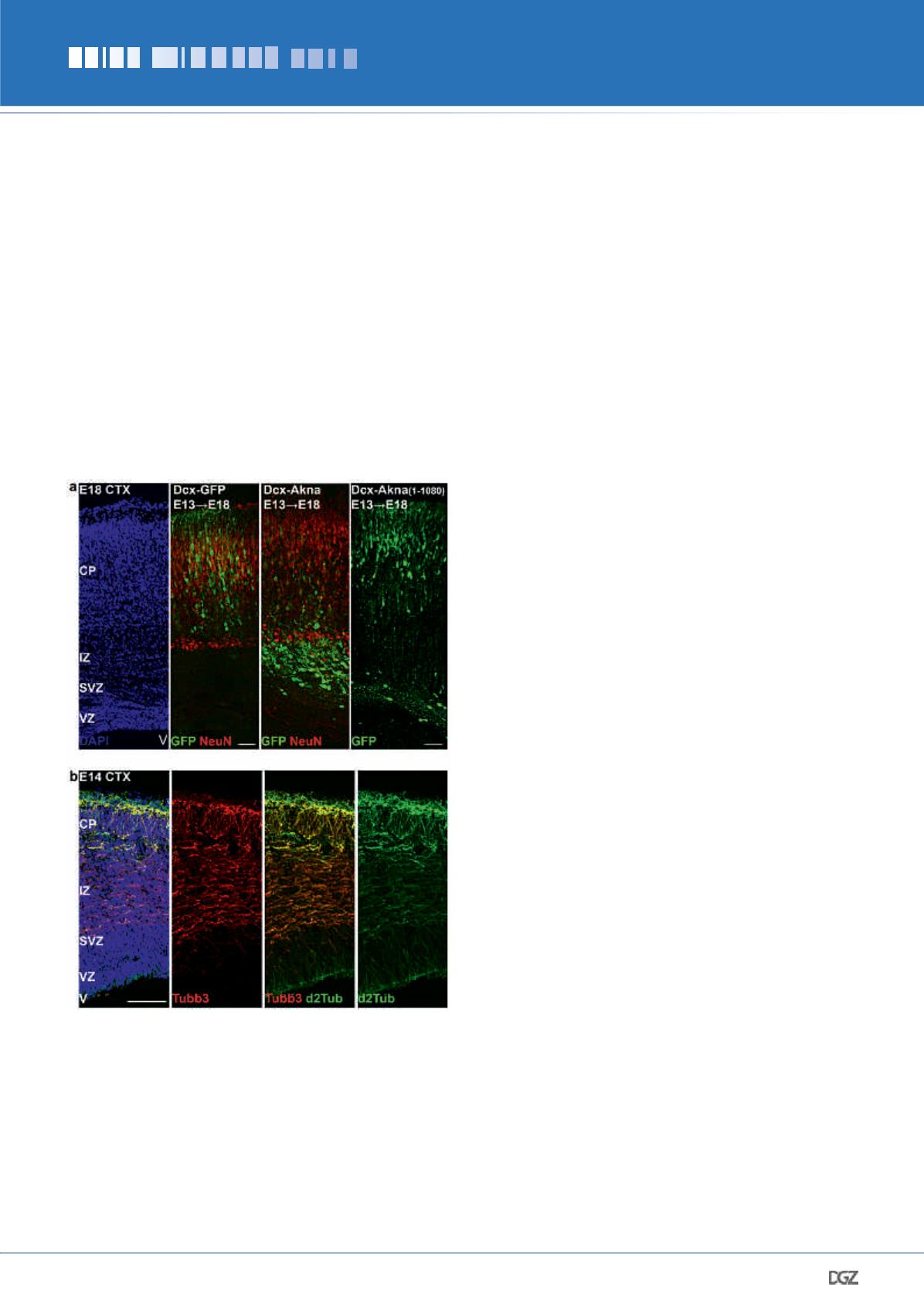
Figure 5:
Akna keeps multipolar BPs in the SVZ. (a) Overexpression of
Akna cDNA under the promoter of the early neuronal gene Doublecortin
(Dcx) impairs the migration of SVZ cells to the CP. Thus, endogenous
downregulation is a requirement to leave the SVZ. The centrosomal
localization of Akna is essential for its function, as demonstrated by
the overexpression of a non-centrosomal construct (1-1080), which has
no effect. (b) Micrograph of a E14 CTX sections showing enrichment of
de-tyrosinated Tubulin (d2Tub) in Tubb3-positive neurons, high highest
levels I the CP. This indicates that the MTs of neurons are less dynamic
than progenitor cells in the SVZ (notice low levels of d2Tub there).
Bars = 50 μm.


