Cell News 04/2019
22
PRIZE WINNERS 2019
The mammalian lymphatic system consists of strategically
located lymph nodes (LNs) embedded into a lymphatic vascular
network. Lymphatic capillaries take up interstitial fluid, antigens
and antigen-presenting cells, and collecting vessels transport
lymph to LNs. Afferent collecting lymphatics deliver lymph to the
LN subcapsular sinus (SCS) through a fibrous capsule surround-
ing these organs. From the SCS, lymph reaches trabecular and
medullary lymphatic sinuses and exits via the efferent collecting
vessels. Such organization is important for intranodal migration
of dendritic cells (Ulvmar et al., 2014), filtration of small versus
large molecular weight components (Rantakari et al., 2015) and
immune tolerance (Cohen et al., 2010; Lund et al., 2012; Tewalt
et al., 2012). However, mechanisms underlying development of
this highly organized system are not fully understood.
Pioneering studies on LN organogenesis have identified key roles
for hematopoietic lymphoid tissue inducer (LTi) and stromal lym-
phoid tissue organizer (LTo) cells, proposing a model where LNs
are initiated after CXCR5
+
pre-LTi cell egress to specific locations
from blood vessels in response to LTo cell-derived CXCL13 (Ansel
et al., 2000; Brendolan and Caamano, 2012; Luther et al., 2003;
Mebius et al., 2001; Ohl et al., 2003; Pavert and Mebius, 2010;
van de Pavert et al., 2009; Yoshida et al., 2001). Clustering and
crosstalk of lymphotoxin-
α
1
β
2 (LT
αβ
)
+
LTi and lymphotoxin ß
receptor (LTßR)
+
LTo cells result in further signal amplification,
leading to LTi cell maturation and accumulation. Recently, the
role of lymphatic and blood vessels in LN initiation and develop-
ment have been under debate (Onder et al., 2017; Vondenhoff et
al., 2009b), and the mechanisms of LN capsule and SCS forma-
tion still remain enigmatic.
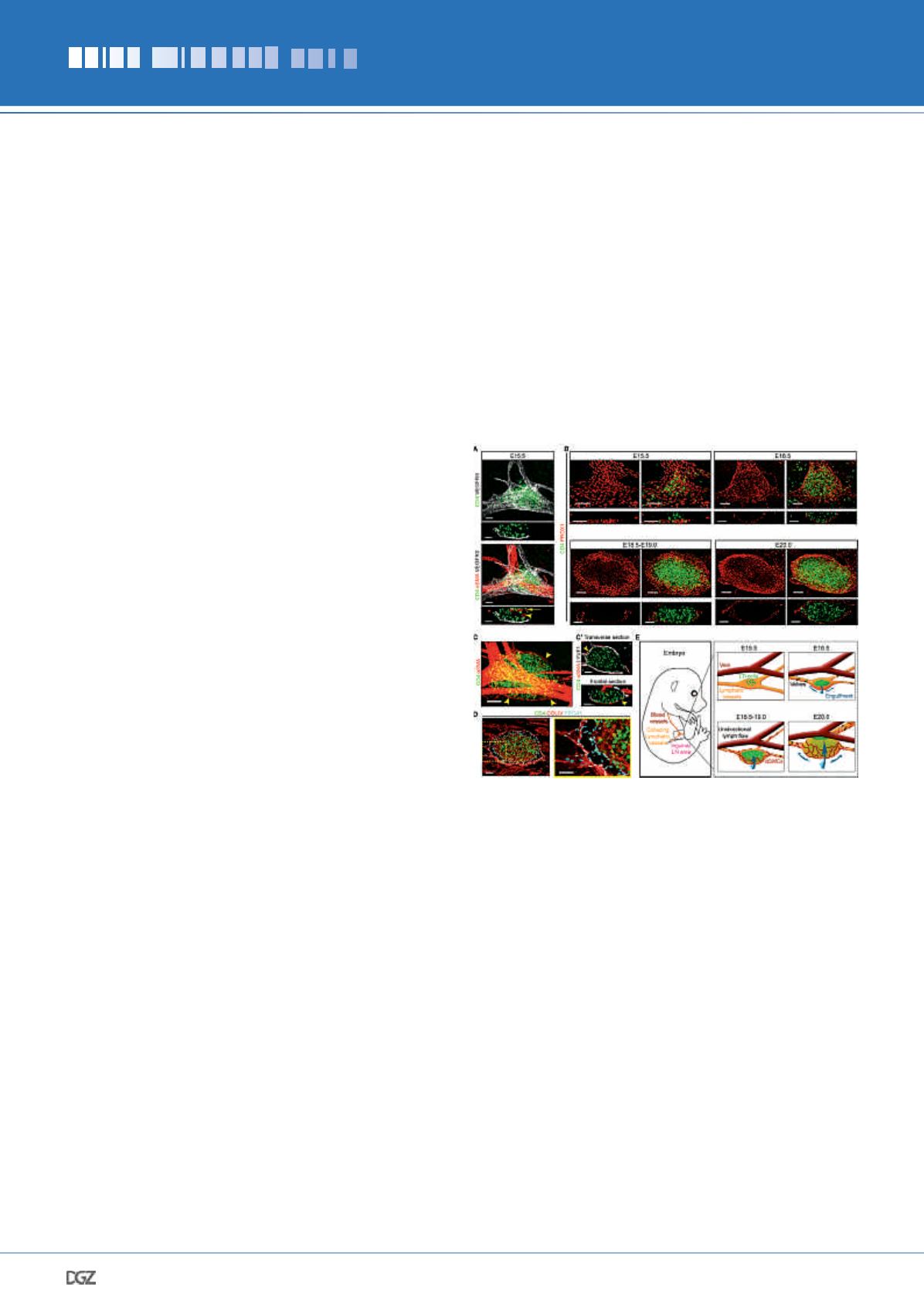
To understand how lymphatic vessels participate in LN develop-
ment and how the SCS forms, we first analyzed embryonic in-
guinal LNs (iLNs) at different stages using whole-mount imaging
(Fig. 1). At E15.5, single extravascular CD4
+
LTi cells and aggre-
gates accumulated at bifurcations of the inguinal blood vessels,
beneath the subepigastric vein/artery and above lymphatic
vessels (Fig. 1, A and B). At E16.5, the lymphatic vessel beneath
the LN anlage expanded and formed a shallow disk (Fig. 1B).
The disk progressively enlarged in a coordinated double-lym-
phatic endothelial cell (LEC) layer, forming a cup-like structure
growing around CD4
+
LTi cell area and almost fully engulfed it
by E20-E20.5 (Fig. 1, B-D). At E18.5, smooth muscle cells (SMCs)
were associated with the exterior LN LECs and closely followed
the expanding LEC sheet (Fig. 1, C and C’), indicating coordi-
nated formation of the LN capsule. Staining for collagen IV also
revealed increased extracellular matrix deposition around the
LN capsule (Fig. 1D). These data demonstrated that the growing
Werner Risau Prize
Multiple roles of lymphatic vessels in
peripheral lymph node development
Esther Bovay
Figure 1. Lymphatic vessel remodeling, SMC recruitment and deposition
of basement membrane during LN development.
(A) LTi cells accumulate between lymphatic vessels and the subepigas-
tric vein. Whole-mount and frontal view (10-μm) of iLN; CD4 (green),
VEGFR3 (white) and
α
SMA (red). Arrowhead, artery; arrow, vein.
E15.5
n
= 6. Scale bar, 50 μm. (B) Lymphatic remodeling during iLN
development. Whole-mount and frontal views (10-μm); PROX1 (red)
and CD4 (green). E15.5
n
= 4; E16.5
n
= 5; E18.5-E19.0
n
= 6; E20.0
n
= 3. Scale bar, 50 μm. (C) SMCs surround iLN. Whole-mount;
α
SMA
(red) and CD4 (green). Arrowheads, LN SMCs. E18.5
n
= 3. Scale bar,
50 μm. (C’) Transverse and frontal views (1-μm) of
C
, LYVE1 (white).
Scale bar, 50 μm. (D) Extracellular matrix deposition around iLN.
Whole-mount 10-μm transverse view; collagen IV (red), PROX1 (blue)
and CD4 (green). Right panel: high-magnification image. E18.5
n
= 3.
Scale bars, 50 μm and 30 μm. (E) LN capsule expansion steps. Artery
and blood capillaries inside the LN are not shown.


